{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Mecinonota regia
sumatrana
Mecinonota regia
sumatrana熱帯林の昆虫相は温帯林などに比べて豊かであるといわれている。Erwin( 1988)が大胆に予想したところでは熱帯アメリカだけで3千万種いるという計算になる 。しかし、実際にどれほどの種類の昆虫が熱帯林にいるのかは、まだだれにもわかっ ていない。これは、昆虫類が種類が多くまだ、記載すらすんでいないものが多い上に 、多様な生活形をとるため、生息場所や発生時期、行動習性などが多様であり、一時 に全ての昆虫を捕獲することなど不可能だからである。さらに、それに追い打ちをか けているのが、熱帯林の樹木の高さである。
典型的な熱帯降雨林では、超高木と呼ばれるものは、50から70mもあるのが普 通である。熱帯林を訪れたことがある方ならば、おわかりになるだろうが、熱帯林に は沢山の昆虫に会えると期待して林内に入ると、期待とはうらはらに、ほとんど虫に 出会うことがない。これは、多くの昆虫類が、最も生産活動が活発に行われている林 冠部におり、我々がいる地表付近にまでは降りてこないためではないかといわれてい る。実際に、ライトトラップを異なる高さに設置したヨコバイ類の調査では、30mの 林冠部分で最も多くとれている(Rees, 1983)。我々は、いってみれば熱帯林という深い海の底をはいずりまわっている貝よ うなもので、多くの昆虫達が泳ぎ回っている?海面に容易には浮び上がることはでき ないのである。
昆虫類の中でも、甲虫類はその種類の多さ(全昆虫の40%)森林のあらゆるハ ビタットを利用している生活形の多様さからいっても重要なグループの一つである。 また、甲虫は成虫が比較的長生きするため、成虫を捕獲できる確率が高い。さらにか らだが頑丈であるため、トラップによる調査でも標本の破損が少なく、その後の処理 が容易である。そして、比較的熱帯での研究がすすんでおらず、熱帯林生態系内での 位置付けがおくれている。そのなかでも特に、熱帯林の動態の上で重要な役割を果た している花粉媒介者としての甲虫(コガネムシ、ハナノミ、ケシキスイ類など)( Kevan & Baker, 1983)と材の分解者としての甲虫(キクイムシ、ゾウムシ、カミキリムシ類)に注目 することにした。
幸い、訪花性の昆虫類に対して有効な誘引トラップが開発されており(Ikeda et al. ,1993)、キクイムシについてもエタノールによってかなり捕獲できることがわかって いた。
しかし、問題は、いかにして林冠にトラップを上げるかであった。
予算の関係からロープを林冠の枝にひっかけてトラップを林冠から吊り下げる 方式が最もよいと思われた。しかし、超高木の枝から吊り下げるのでは、林冠の上に 設置することができないため、森林内での訪花性甲虫の垂直的な活動分布などを完全 にはおさえられない。そこで、気球を使ってトラップを吊り下げる方法を試みること にした。これなら、林冠の上にまでトラップを上げることができそうである。
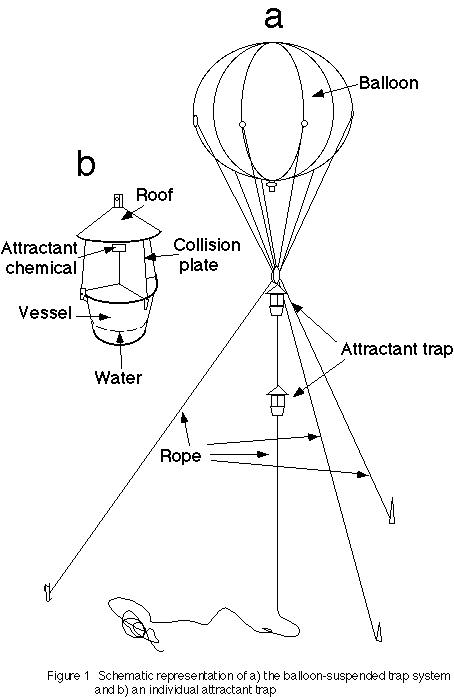
バルーントラップシステムとは、広告用の気球を用いてトラップを林冠の上部 まで上げる方法である(図-1)(Fukuyama et al., 1994)。使用するトラップはあまり重くないもの(4kg以下)ならどんなものでもよ い。今回は、コガネムシやカミキリムシなどの甲虫を捕獲するために開発されたサン ケイ化学社製の衝突板式トラップ(図-1b)(写真1)を用いた。
このトラップは雨よけの屋根、衝突板と捕獲容器からできており、交差した衝 突板の中央に誘引剤をおくようになっている。衝突して落下した昆虫を捕獲するため に、衝突板の下に容器があり、界面活性剤と防腐剤(ソルビン酸)を入れた水が入れ てある。主に訪花性甲虫類を採集するために、白色トラップを用い、誘引剤としては 花の匂い成分の一種であるベンジルアセテートを用いた。また、キクイムシ類を採集 するために黒色トラップを用い、誘引剤はエタノールを用いた。ベンジルアセテート はもともと訪花性カミキリムシのスギノアカネトラカミキリ用に開発された誘 引剤であるが、他の多くの訪花性甲虫類を効率よく採集できるといわれている(槙原 私信)。
バルーンは白色の塩化ビニール製フィルム(厚さ0.12mm)でできた直径2.5mの 球形をしており、重量はロープなどを含め約3kgである。7m3のヘリウム ガスを充填すると約4kgの揚力が得られる。ちなみに、今回用いたトラップは総重量 が約1.5kgであるので、2器までつり下げることが可能である。バルーンは安定させる ために3本の細いロープで係留する( 図-1a)。
国内での予備実験
果たして、こんな方法でトラップを林冠にどのくらいの期間あげておくことが できるか、風の影響をどの程度受けるのか、安定性はどうか、破損がどの程度発生す るかを調べるため、まず国内で予備実験を行った。試験地は札幌市羊ケ丘の森林総合 研究所の実験林(広葉樹林)である。その結果、林外の瞬間最大風速が 24.12 km /h であっても、林内ならば継続して係留が可能であり、トラップに訪花性昆虫が捕獲さ れた(写真2)。ただし、バルーンはかなり移動し、林冠に 当たることが多く、トラップもかなり揺れ動いたため実際の調査には適さないと思わ れた。しかし、ロープを3本にするとかなり安定することがわかった。
熱帯林での実験
比較的風の強い北海道でも、継続係留が可能であったことから、熱帯林のなか でもモンスーンの影響を受けない半島マレーシアの熱帯降雨林ならば、年間にわたっ て風が弱いので、調査ができそうであるという目途が立った。そこで今度は実際に熱 帯降雨林で試すことにした。
調査地は、半島マレーシアのパソーにあるフタバガキ科の高木が優占する低地 熱帯降雨林の保護林に設定した。林の入り口から約800m入った地点に互いに100mほ ど離れた2カ所の調査地点(A、B)を設定し、各地点にバルーンを1つづつあげた( 写真3)(写真4)。両地 点の最高樹高は約30mであった。
バルーンには、原則として2器のトラップをとりつけ、高さやトラップの種類を 変えながら10日間調査を行った。トラップの高さは1.5m(地表付近)、7m(林冠下 部)、15(林冠中部)、21、25m(林冠上部)としトラップの種類は白色衝突版にベ ンジルアセテートと黒色衝突版にエタノールの組み合わせの2種類とした。なお、高 さ21mはB地点のみ、25mについてはA地点のみでそれぞれ1日しか実施しなかった。 調査は1992年1月14日から25日まで行い、回収は中間の3日間を除き原則として毎日行 った。
熱帯林でのバルーントラップの係留状況
調査期間中、いずれの高さにおいてもバルーンは安定して係留され、トラップ もほとんど振動しなかった(写真5)。心配された風の影響 はほとんどなく、午後のスコールの時を除けば無風状態であった。調査期間中の最大 風速は14.4 km/hであり、この地域の年間の最大風速が14.4 km/hを越えるのは10日前後しかないこと、最大でも19 km/hにしかならないことなどから、この地域においては年間を通じて調査が可能であ ると思われる。また、ガスの漏洩もわずかで10日間継続して係留することができた。 ただし、当初B地点でロープ1本による係留を試したところ、バルーンがかなり振動 し樹木の枝にロープがからみつくというアクシデントがあった。からみついたバルー ンを引き下ろす課程でバルーンが破損し初めの1週間はB地点では係留調査ができな かった。3人(少なくとも2人)いればバルーンの設置が行えトラップの回収のため の上げ下げは1人でも容易にできることがわかった。
トラップに誘引捕獲された甲虫類
調査期間中に合わせて13科(1上科を含む)の甲虫類が捕獲された。最も個体数 が多かったのがScarabaeidae(コガネムシ科) で約64%を占めた。次いTrogossitidae(コクヌスト科)、Curculionidae(ゾウムシ科)、Carabidae(オサムシ科)、 Chrysomelidae(ハムシ科)、Staphylinoidea(ハネカクシ上科)、Elateridae(コ メツキムシ科)、Mordellidae(ハナノミ科)、 Nitidulidae(ケシキスイ科)、Anthribidae(ヒゲナガゾウムシ科)、Erotylidae( オオキノコムシ科)、Cerambycidae(カミキリムシ科)、Scolytidae(キクイムシ科)の順であり、主な訪花性甲虫 のグループ(Kevan& Baker, 1983)は捕獲できた。
ベンジルアセテートに誘引されたものとしてはコガネムシ科のハナムグリ類 Dasyvalgus3種、Mecinonota 1種が優占しており(写真-2,写真-3) Xylosandrus crassiusculusが優占した(写真-4)。
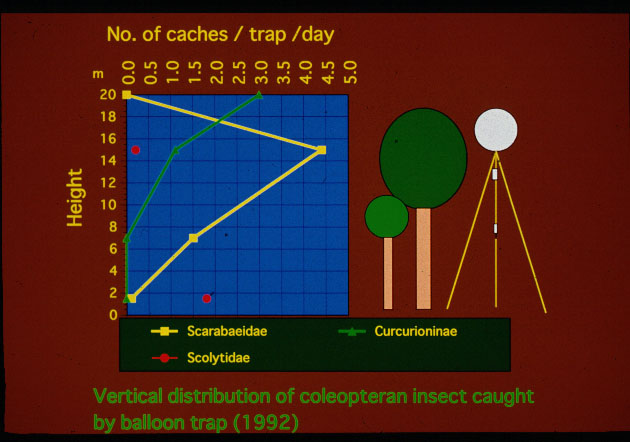
高さによる捕獲傾向の違いをみると、全体では明らかにトラップをあげた地点 での林冠中層にあたる15m付近でもっとも多く、地表付近では、きわめて少なかった (図-3)。ただし、この傾向は種類によって異なり、最優占種 であるDasyvalgus 3種は林冠中層に多く地表付近ではまったく捕獲されなかったのに対して同じコ ガネムシ科のMecinonota の1種は地表付近でも捕獲されている(Table1)。また 、花や新梢、葉などを食べていると思われる(森本私信)Endaeus 2種は、林冠の上部で最も多くとれている(図-3のゾウムシ類)。2つの調査地点の間 では、捕獲された甲虫の種類組成はほとんど変わらないが、Endaeus属のゾウ ムシはB地点の方が明らかに多かった(Table1)。Endaeusが林冠表層に集中 して活動していると仮定すると、トラップを設置した場所の林冠の立体的な構造の違 いが敏感に反映されているのかもしれない。
気球を使って林冠の昆虫を調査するという一見、突飛な方法も、熱帯林のよう な風の少ないところでは、十分可能であり有用な方法であることがわかった。この方 法の、利点は、タワーやウオークウエイのように場所が限定されることがないため、 対象とする樹木などの近くに直接トラップを上げることができることであろう。その ため、そのとき開花している樹木を対象に訪花昆虫をを調査したり、林縁から林内に かけての群集の変化を解析したりでき、熱帯林の昆虫群集を水平的および垂直的に調 査ができる。特に、コガネムシ類のように土壌表層に生息しているものが一時的に林 冠を訪れるものを短期間で調査するのには適している。
さらに、通常の樹木に懸垂させる方法に比べて、林内の小さなギャップに上げ たり、林冠の上部にまで上げることができ、トラップの選択によって、かなりきめ細 かい調査も可能となる。また、飛行船やタワークレーンのように多大な経費も必要で はなく(気球1個当たり7万円。ガス代が1回に約2万円)、要員もわずかですみ、森林 環境にもほとんど影響をあたえることがなく、これまでの方法に比べコストパフォー マンスがよいと思われる。
欠点としては、気球であるため、風の影響に弱いことと、薄いビニール製であ るためわずかなことで破損することがあるため、長期間のメンテナンスなしの継続使 用にはむかないこと。また、直接観察や採集ができないので、より高度な生態調査な どには利用できないことやトラップに制限を受けるため特定の昆虫群集にしか利用で きない点などである。ただし、ビデオカメラなどを係留することは可能なので、不十 分ながら間接的な観察は可能であるし、ライトトラップやマレーズトラップのような かなり広範囲の昆虫をあつめるトラップを利用することもできる。また、トラップの 改良によりマーク再捕獲法などにより、成虫の移動などの調査にも応用が期待できる と思われる。
Erwin, T.L. 1988. The tropical forest canopy-the heart of biotic diversity. In Biodiversity, Wilson, E.O. ed., pp123-129, National Academy Press, Washington, D.C.
福山研二(1995)いかにして熱帯林の林冠部での甲虫類の活動を探るか-半島マ レーシア・パソーでの試み-, TROPICS 4(4):317-326.
Ikeda T., Ohya E., Makihara H., Nakashima T., Saito T., Tate T. & Kojima K. (1993) Olfactory responses of Anaglyptus subfasciatus Pic and Demonax transilis Bates (Coleoptera: Cerambycidae) to flower scents. Journal of the Japanese Forestry Society. 75(2):108-112.
Kevan, P.G. & Baker, H.G. 1983. Insects as Flower Visitors and Pollinatiors,. Ann.
Rev. Entomol. 28:407-453.
Rees, C.J.C. 1983. Microclimate and the flying Hemiptera fauna of a primary lowland rain forest in Sulawesi. In Tropical Rain Forest: Ecology and Management, Sutton, S.L. et al. eds., 121-136, Blackwell Sci. Publ., Oxford.
Table 1. The number of flower visiting beetles collected using benzyl acetate as an attractant in traps placed simultaneously at different heights or in different sites over consecutive days.
(Species Traps at different heights Taps in different sites in site A at 15 m )
|
|
|
|
|
|
|
|
|
|
Macronota sp. |
|
|
|
|
|
|
|
|
Dasyvalgus sp.1 |
|
|
|
|
|
|
|
|
D. sp.2 |
|
|
|
|
|
|
|
|
D. sp.3 |
|
|
|
|
|
|
|
|
Endaeus sp.1 |
|
|
|
|
|
|
|
|
E. sp.2 |
|
|
|
|
|
|
|
|
Total individuals |
|
|
|
|
|
|
|
|
Total species |
|
|
|
|
|
|
|
Mecinonota regia
sumatrana
 Dasyvalgus
vethii
Dasyvalgus
vethii
 D. niger
D. niger
 D.
striatippennis
D.
striatippennis
 D.
sumatranus
D.
sumatranus
 D.
biguttatus
D.
biguttatus
 D.
setipygus
D.
setipygus
 D.
dohli
D.
dohli